![(A) Young yellow-orange pseudoflower. (B) Mature pseudoflower. (C) Longitudinal section of an infected X. surinamensis inflorescence. (D) Healthy yellow flower of X. surinamensis shown for comparison. [SOURCE]](https://images.squarespace-cdn.com/content/v1/544591e6e4b0135285aeb5b6/1612389646771-T82PTY66AT9ULQO2VTPW/1-s2.0-S1087184520301572-gr1.jpg)
(A) Young yellow-orange pseudoflower. (B) Mature pseudoflower. (C) Longitudinal section of an infected X. surinamensis inflorescence. (D) Healthy yellow flower of X. surinamensis shown for comparison. [SOURCE]
Imagine there was a fungus that was able to hijack human reproductive structures so that it could reproduce. Though this sounds like the basis of a strange science fiction story, a similar situation to this has just been described from Guyana between two species of yellow-eyed grass (Xyris setigera & X. surinamensis) and a newly described species of fungus called Fusarium xyrophilum.
Fungi that hijack plant reproductive systems are pretty rare in nature, especially when you consider the breadth of interactions between these two branches on the tree of life. What makes this newly described case of floral hijacking so remarkable is the complexity of the whole process. It all begins when an infected Xyris host begins to produce its characteristically lanky inflorescence.
At first glance, nothing would appear abnormal. The floral spike elongates and the inflorescence at the tip gradually matures until the flowers are ready to open. Even when the “flower” begins to emerge from between the tightly packed bracts the process seems pretty par for the course. Gradually a bright yellow, flower-like structure bursts forth, looking very much like how a bright yellow Xyris flower should look. However, a closer inspection of an infected plant would reveal something very different indeed.
Instead of petals, anthers, and a pistil, infected inflorescences produce what is called a pseudoflower complete with petal-like structures. This pseudoflower is not botanical at all. It is made entirely by the Fusarium fungus. Amazingly, these similarities are far from superficial. When researchers analyzed these pseudoflowers, they found that they are extremely close mimics of an actual Xyris flower in more than just looks. For starters, they produce pigments that reflect UV light in much the same way that actual flowers do. They also emit a complex suite of volatile scent compounds that are known to attract pollinating insects. In fact, at least one of those compounds was an exact match to a scent compound produced by the flowers of these two Xyris species.
So, why would a fungus go through all the trouble of mimicking its hosts flowers so accurately? For sex, of course! This species of Fusarium cannot exist without its Xyris hosts. However, Xyris don’t live forever and for the cycle to continue, Fusarium must go on to infect other Xyris individuals. This is where those pseudoflowers come in. Because they so closely match actual Xyris flowers in both appearance and smell, pollinating bees treat them just like flowers. The bees land on and investigate the fungal structure until they figure out there is no reward. No matter, they have already been covered in Fusarium spores.
As the bees visit other Xyris plants in the area, they inevitably deposit spores onto each plant they land on. Essentially, they are being coopted by the fungus in order to find new hosts. By mimicking flowers, the Fusarium is able to hijack plant-pollinator interactions for its own reproduction. It is not entirely certain at this point just how specific this fungus is to these two Xyris species. A search for other potential hosts turned up only a single case of it infecting another Xyris. It is also uncertain as to how much of an impact this fungus has on Xyris reproduction. Though the fungus effectively sterilizes its host, researchers did make a point to mention that Xyris populations may actually benefit from having a few infected plants as the pseudoflowers last much longer than the actual flowers and therefore could serve to attract more pollinators to the area over time. Who knows what further investigations into the ecology of this bizarre system will reveal.
Photo Credit: [1]
Further Reading: [1]





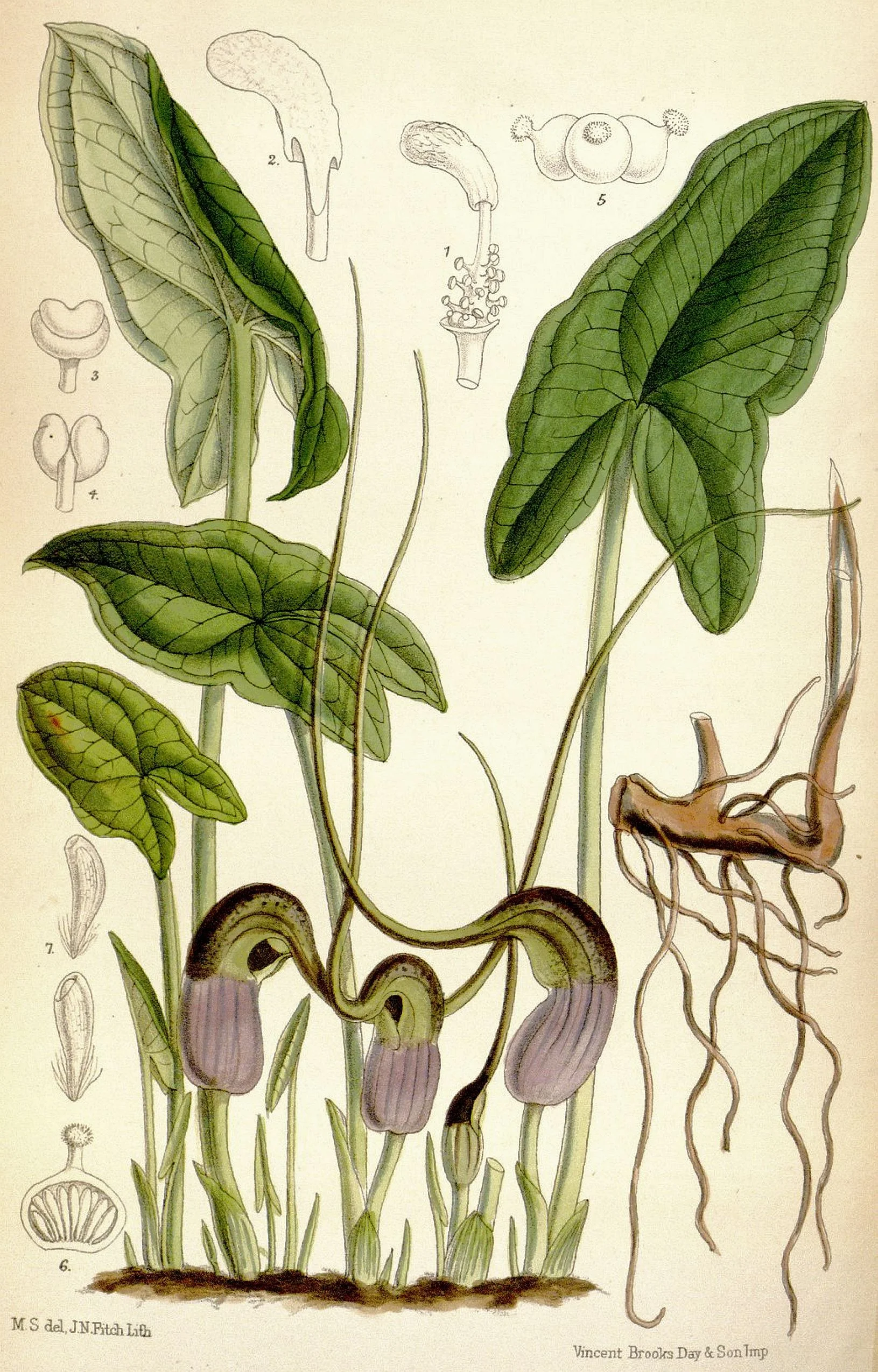
![Anatomy of a mouse plant inflorescence [SOURCE]](https://images.squarespace-cdn.com/content/v1/544591e6e4b0135285aeb5b6/1557945730300-PS22LHE82GUKE9OWKEJA/mouse+cross.JPG)



![Stems climbing on fallen dead wood (a) or on standing living trees (b). A thick and densely branched root clump (c) and thin and elongate roots (d) [Source]](https://images.squarespace-cdn.com/content/v1/544591e6e4b0135285aeb5b6/1518471929195-C99KP24U24J238YMX055/myco+roots.JPG)


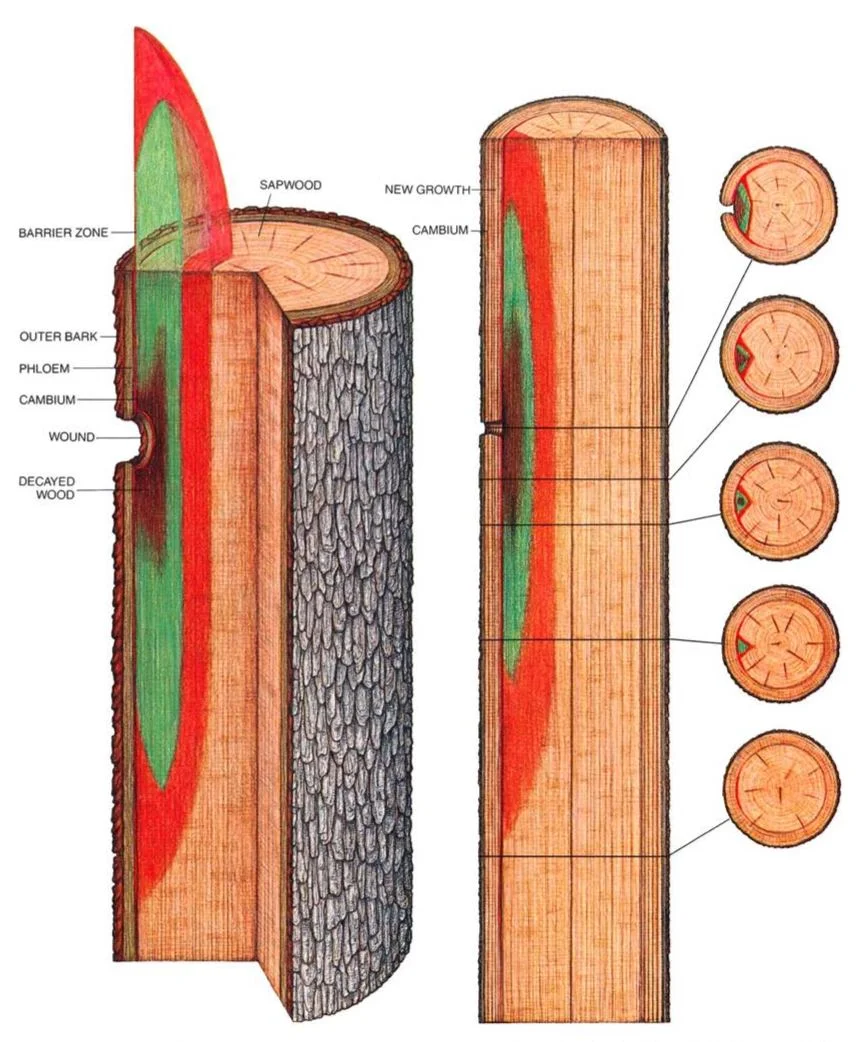




![[SOURCE]](https://images.squarespace-cdn.com/content/v1/544591e6e4b0135285aeb5b6/1485992214448-W3DN42037E3R1Z3OYFIJ/image-asset.jpeg)







