Reconstructing extinct organisms based on fossils is no simple task. Rarely do paleontologists find complete specimens. More often, reconstructions are based on fragments of individuals found either near one another or at least in similar rock formations. This is especially true for plants as their growth habits frequently result in fragmentary fossilization. As such, fossilized plant remains of a single species are often described as distinct species until subsequent detective work pieces together a more complete picture.
Such was the case for the fossil remains of what were described as Krassilovia mongolica and Podozamites harrisii. Hailing from the Early Cretaceous (some 100-120 million years ago), Krassilovia was only known from oddly spiny cone scales and Podozamites was only known from strap-shaped leaves found in a remote region of Mongolia. Little evidence existed to suggest they belonged to the same plant. That is, until these structures were analyzed using scanning electron micrographs.

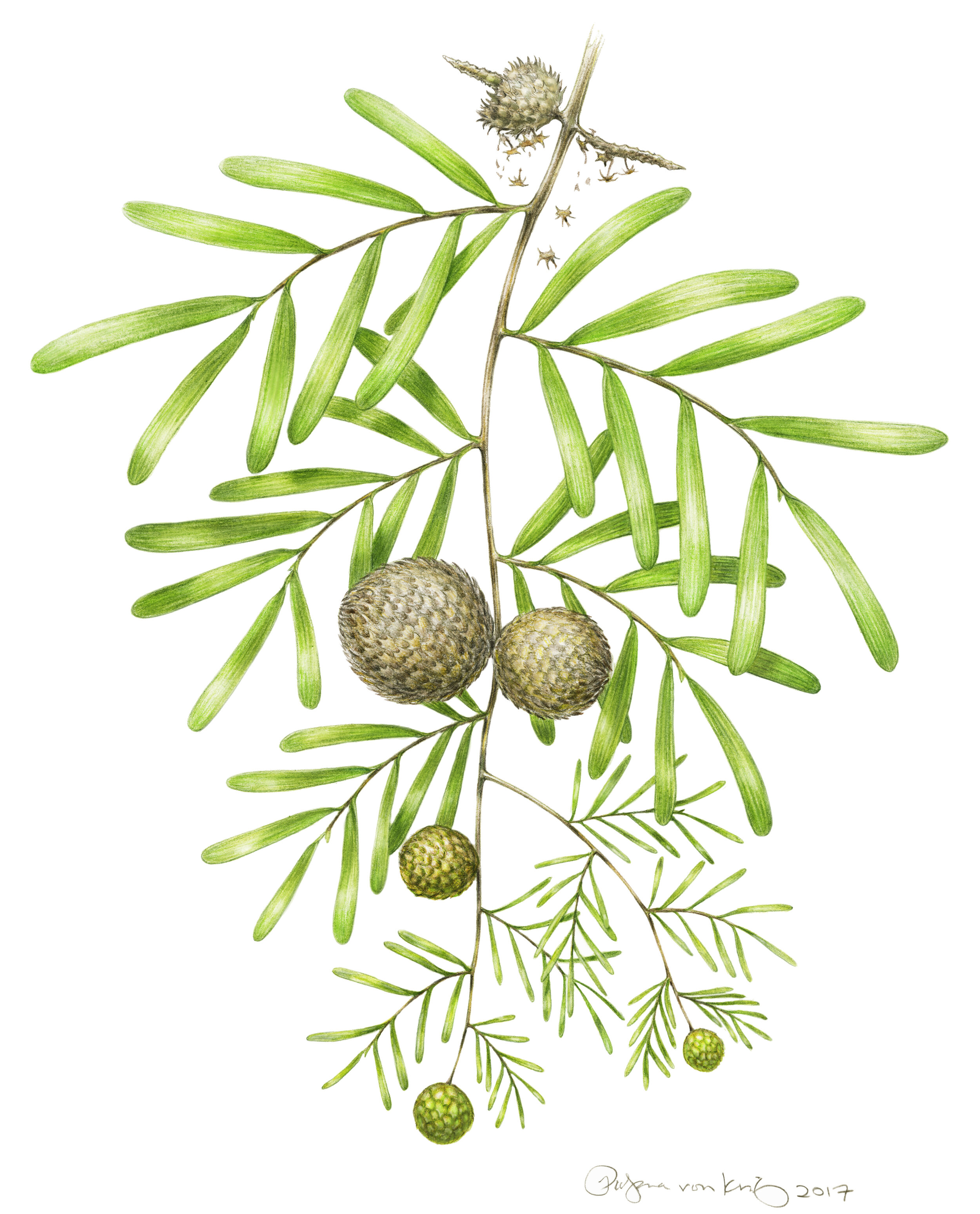
(A–C) Articulated seed cones, (D) Isolated cone axis, (E) Incomplete leafy shoot showing a cluster of three attached leaves, (F) Three detached strap-shaped leaves, G) Detail of A showing tightly imbricate interlocking bract-scale complexes, (H) Detail of leaf apex showing converging veins, (I) Three isolated bract-scale complexes showing abaxial (top) and adaxial (bottom) surfaces, (J) Two isolated seeds showing narrow wings. [SOURCE]
These fossilized plant remains were preserved in such detail that microscopic anatomical features such as stomata were visible under magnification. By studying the remains of these plants as well as others, scientists discovered some amazing similarities in the stomata of Krassilovia and Podozamites. Unlike other plant remains associated with those formations, the Krassilovia cone scales and Podozamites leaves shared the exact same stomate morphology. Though not without some uncertainty, the odds that these two associated structures would share this unique morphological trait by chance is slim and suggests that these are indeed parts of the same plant.
The amazing discoveries do not end with stomata either. After countless hours of searching, fully articulated Krassilovia cones were eventually discovered, which finally put the strange spiky cone scales into context. It turns out those spiked scales interlocked with one another, with the two bottom spikes of one scale interlocking with the three top spikes of the scale below it. In life, such interlocking may have helped protect the developing seeds within until they had matured enough to be released. Also, the sheer volume of cone scales coupled with other minute anatomical details I won’t go into here indicate that, similar to Abies and Cedrus cones, Krassilovia cones completely fell apart when fully ripe.

Though not related, the cone scales of the extinct Krassilovia (left) show similarities with the cone scales of modern day Cryptomeria species (right).
Interestingly, the ability to resolve microscopic structures in these fossils has also provided insights into some modern day taxonomic confusion. It turns out that Krassilovia shares many minute anatomical similarities with present day Gnetales. Gnetales really challenge our perception of gymnosperms and their superficial resemblance to angiosperms have led many to suggest that they represent a clade that is sister to flowering plants. However, more recent molecular work has placed the extant members of Gnetales as sister to the pines. Evidence of shared morphological features between extinct conifers like Krassilovia and modern day Gnetales add some interesting support to this hypothesis. Until more concrete evidence is described and analyzed, the true evolutionary relationships among these groups will remain the object of heated debate for the foreseeable fture.
What we can say is that Krassilovia mongolica was one remarkable conifer. Its unique morphology clearly demonstrates that conifers were once far more diverse in form and function than they are currently. Even the habitat in which Krassilovia once lived is not the kind of place you can find thriving conifer communities today. Krassilovia once grew in a swampy habitat. However, whereas only a few extant conifers enjoy swamps, Krassilovia once shared its habitat with a wide variety of conifer species, the likes of which we are only just beginning to appreciate. I for one am extremely excited to see what new fossil discoveries will uncover in the future.
Further Reading: [1]